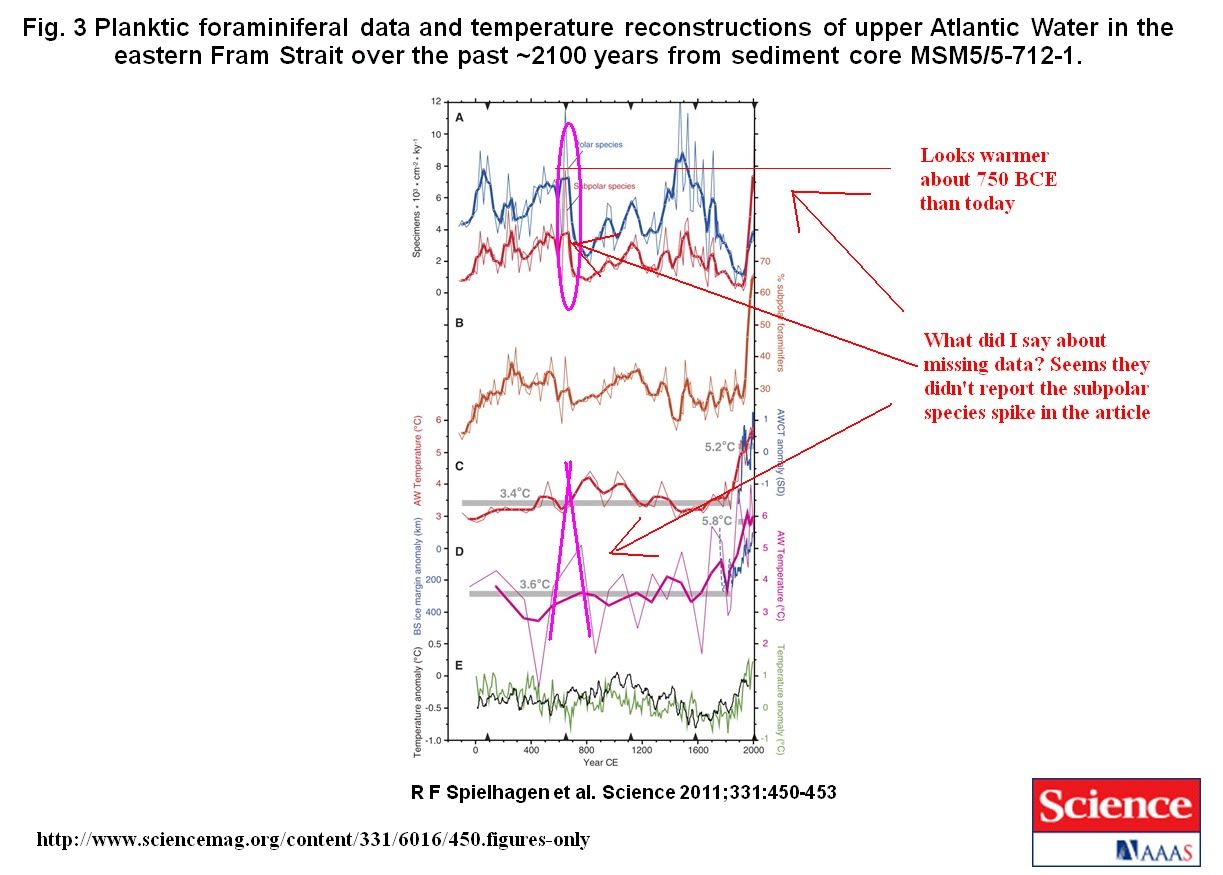
For Mg/Ca analysis ~50 tests of N. pachyderma (sin.) per sample were picked from the 100-150 μm fraction. The size interval was kept narrow in order to avoid a possible size dependent bias (S6). In preparation of Mg/Ca measurements, foraminifers were gently crushed between glass slides. Subsequently each sample underwent the trace metal cleaning procedures described elsewhere (S7, S8). Samples were analyzed for Mg/Ca by magnetic sector single-collector ICP-MS on a Thermo-Finnigan Element2, using methods adapted from Rosenthal et al. (S9) with a long-term 1σ precision of Mg/Ca for of 0.5% (S10). Four samples were rejected due to low post-cleaning mass recovery (< 5μg CaCO3) (S10), leaving a total of 26 reliable Mg/Ca data (including two replicates). Elemental ratios of Fe/Ca and Al/Ca (detrital material) and Mn/Ca (secondary diagenetic coatings) were analyzed coincidental with Mg/Ca. No values of Mn/Ca exceeded 24 μmol mol-1, which is well below the threshold for likely trace metal contamination (S11). One sample (0.25 cm) had Fe/Ca and Al/Ca values exceeding 100 μmol mol-1. However, this single sample was not omitted as its Mn/Ca ratio did not show signs of contamination and its Mg/Ca value was similar to surrounding samples.
To convert the measured Mg/Ca ratios into temperatures we used the N. pachyderma equation of Elderfield and Ganssen (S12): Mg/Ca (mmol mol-1) = 0.5 exp 0.10 T. This equation is indistinguishable from the Norwegian Sea N. pachyderma (sin.) calibration of Nürnberg (S13)
and the cultured N. pachyderma (dex.) calibration of von Langen et al. (S14). Application of an alternative equation of Kozdon et al. (S15) results in consistently lower temperatures throughout the core which are also lower than the SIMMAX results. Tests from the upper 10 cm of the core showed some signs of dissolution and were fragile. Generally, partly dissolved tests should be avoided as they may give too low Mg/Ca ratios and the reconstructed temperatures may be underestimated (S16). With respect to core MSM5/5-712-1, however, we regard the close similarity between temperatures reconstructed from independent SIMMAX and Mg/Ca results as evidence of a minor effect of dissolution.
Were dissolution of importance, it would affect also the planktic foraminifer associations used
for SIMMAX. Fragile subpolar species would be more easily dissolved than the only polar, robust species N. pachyderma (sin.), eventually leading to lower calculated SIMMAX temperatures especially for the youngest time interval. Thus, carbonate dissolution within the uppermost part of the core would result in an underestimation (by both methods) of the Atlantic Water temperature difference between the Modern (Industrial) Period and, e.g., the Medieval Climate Anomaly.
Replicate Mg/Ca analysis on separate picks at 0 cm and 28 cm core depth revealed differences of 0.01 mmol mol-1 and 0.12 mmol mol-1, respectively. Although this is insufficient to provide a robust estimate of reproducibility, a pooled standard deviation of order 0.05-0.1 mmol mol-1 is typical for foraminifera (e.g., Refs. S17, S18). The standard deviation for each interval before and after AD 1850 was 0.1 mmol mol-1, which translates to relatively large temperature ranges at the cold end of the Elderfield and Ganssen (S12) exponential calibration: +1.3/-1.5C pre-industrial and +1.1/-1.2C post-industrial. Since this is close to our estimated reproducibility, we cannot interpret the temperature variability within each interval, which we suggest is dominated by random error. However the mean within each interval is well constrained, with standard errors (stdev/sqrt[n]) of ±0.3C pre-industrial and ±0.5C post-industrial.